
|
|
|
|
 Far Far |
| WinNavigator |
| Frigate |
| Norton
Commander |
| WinNC |
| Dos
Navigator |
| Servant
Salamander |
| Turbo
Browser |
|
|
| Winamp,
Skins, Plugins |
| Необходимые
Утилиты |
| Текстовые
редакторы |
| Юмор |
|
|

|
File managers and best utilites |
Понятие, история открытия, структура и роль комплекса Гольджи. Из одной клетки состоит клоп аппарат гольджи
Глава 15. Аппарат (комплекс) Гольджи
В 1898 г. итальянский ученый К. Гольджи, используя свойства связывания тяжелых металлов (осмия и серебра) с клеточными структурами, выявил в нервных клетках сетчатые образования, которые он назвал “внутренним сетчатым аппаратом” (рис. 174). Дальнейшее усовершенствование метода окраски металлами (импрегнации) дало возможность убедиться, что сетчатые структуры (аппарат Гольджи) встречаются во всех клетках любых эукариотных организмов. Обычно элементы аппарата Гольджи расположены около ядра, вблизи клеточного центра (центриоли). Участки аппарата Гольджи, четко выявляемые методом импрегнации, имели в некоторых клетках вид сложных сетей, где ячейки были связаны друг с другом или представлялись в виде отдельных темных участков, лежащих независимо друг от друга (диктиосомы), имеющих вид палочек, зерен, вогнутых дисков и т.д. (рис. 175). Между сетчатой и диффузной формой аппарата Гольджи нет принципиального различия, так как часто в одних и тех же клетках наблюдается смена форм этого органоида. Элементы аппарата Гольджи часто связаны с вакуолями, что особенно характерно для секретирующих клеток.
Было обнаружено, что морфология АГ меняется в зависимости от стадий клеточной секреции, что послужило основанием Д.Н. Насонову (1924) выдвинуть гипотезу о том, что АГ является органоидом, обеспечивающим сепарацию и накопление веществ в самых различных клетках.
Долгое время в растительных клетках не удавалось обнаружить элементов аппарата Гольджи обычными методами микротехники. Однако с появлением метода электронной микроскопии элементы АГ были обнаружены во всех растительных клетках, где они расположены по периферии клетки.
Тонкое строение аппарата Гольджи
В электронном микроскопе видно, что аппарат Гольджи представлен мембранными структурами, собранными вместе в небольшой зоне (рис. 176, 177). Отдельная зона скопления этих мембран является диктиосомой (рис. 178). В диктиосоме плотно друг к другу (на расстоянии 20-25 нм) расположены в виде стопки плоские мембранные мешки, или цистерны, между которыми располагаются тонкие прослойки гиалоплазмы. Каждая отдельная цистерна имеет диаметр около 1 мкм и переменную толщину; в центре ее мембраны могут быть сближены (25 нм), а на периферии иметь расширения, ампулы, ширина которых непостоянна. Количество таких мешков в стопке обычно не превышает 5-10. У некоторых одноклеточных их число может достигать 20 штук. Кроме плотно расположенных плоских цистерн в зоне АГ наблюдается множество вакуолей. Мелкие вакуоли встречаются главным образом в периферических участках зоны АГ; иногда видно, как они отшнуровываются от ампулярных расширений на краях плоских цистерн. Принято различать в зоне диктиосомы проксимальный или формирующийся, цис-участок, и дистальный или зрелый, транс-участок (рис. 178). Между ними располагается средний или промежуточный участок АГ.
Во время деления клеток сетчатые формы АГ распадаются до диктиосом, которые пассивно и случайно распределяются по дочерним клеткам. При росте клеток общее количество диктиосом увеличивается.
В секретирующих клетках обычно АГ поляризован: его проксимальная часть обращена к цитоплазме и ядру, а дистальная - к поверхности клетки. В проксимальном участке к стопкам сближенных цистерн примыкает зона мелких гладких пузырьков и коротких мембранных цистерн. В образцах препаративно выделенных зон АГ при негативном контрастировании видно, что к проксимальной части диктиосомы примыкает сетевидная или губкообразная система мембранных полостей. Считается, что эта система может представлять собой зону перехода элементов ЭР в зону аппарата Гольджи (рис. 179).
В средней части диктиосомы периферия каждой цистерны также сопровождается массой мелких вакуолей около 50 нм в диаметре.
В дистальном или транс-участке диктиосом к последней мембранной плоской цистерне примыкает участок, состоящий из трубчатых элементов и массой мелких вакуолей, часто имеющих фибриллярную опушенность по поверхности со стороны цитоплазмы - это опушенные или окаймленные пузырьки такого же типа, как и окаймленные пузырьки при пиноцитозе. Это - так называемая транс-сеть аппарата Гольджи (TGN), где происходит разделение и сортировка секретируемых продуктов. Еще дистальнее располагается группа более крупных вакуолей - это уже продукт слияния мелких вакуолей и образования секреторных вакуолей.
При изучении толстых срезов клеток в мегавольтный электронный микроскоп было найдено, что в клетках отдельные диктосомы могут быть связаны друг с другом системой вакуолей и цистерн. Так что образуется рыхлая трехмерная сеть, выявляемая в световом микроскопе. В случае диффузной формы АГ каждый отдельный его участок представлен диктиосомой. У клеток растений преобладает диффузный тип организации АГ, обычно в среднем на клетку приходится около 20 диктиосом. В клетках животных часто с зоной мембран аппарата Гольджи ассоциированы центриоли; между радиально отходящих от них пучков микротрубочек лежат группы стопок мембран и вакуолей, которые концентрически окружают клеточный центр. Эта связь, вероятно, отражает участие микротрубочек в движении вакуолей.
Секреторная функция аппарата Гольджи
Мембранные элементы АГ участвуют в сегрегации и накоплении продуктов, синтезированных в ЭР, участвуют в их химических перестройках, созревании: это, главным образом перестройка олигосахаридных компонентов гликопротеинов в составе водорастворимых секретов или в составе мембран (рис. 180).
В цистернах АГ происходит синтез полисахаридов, их взаимосвязь с белками, приводящая к образованию мукопротеидов. Но главное, с помощью элементов аппарата Гольджи происходит процесс выведения готовых секретов за пределы клетки. Кроме того, АГ является источником клеточных лизосом.
Участие АГ в процессах выведения секреторных продуктов было очень хорошо изучено на примере экзокринных клеток поджелудочной железы. Для этих клеток характерно наличие большого числа секреторных гранул (зимогеновых гранул), которые представляют собой мембранные пузырьки, заполненные белковым содержимым. В составе белков зимогеновых гранул входят разнообразные ферменты: протеазы, липазы, карбогидразы, нуклеазы. При секреции содержимое этих зимогеновых гранул выбрасывается из клеток в просвет железы, а затем перетекает в полость кишечника. Так как основным продуктом, выводимым клетками поджелудочной железы, является белок, то исследовали последовательность включения радиоактивных аминокислот в различные участки клетки (рис. 181). Для этого животным вводили меченную тритием аминокислоту (3Н-лейцин) и с помощью электронно-микроскопической радиоавтографии следили во времени за локализацией метки. Оказалось, что через короткий промежуток времени (3-5 мин) метка локализовалась только в базальных участках клеток, в участка, богатых гранулярным ЭР. Так как метка включалась в белковую цепь во время синтеза белка, то было ясно, что ни в зоне АГ, ни в самих зимогеновых гранулах синтез белка не происходит, а он синтезируется исключительно в эргастоплазме на рибосомах. Несколько позднее (через 20-40 мин) метка кроме эргастоплазмы была обнаружена в зоне вакуолей АГ. Следовательно, после синтеза в эргастоплазме белок был транспортирован в зону АГ. Еще позднее (через 60 мин) метка обнаруживалась уже и в зоне зимогеновых гранул. В дальнейшем метку можно было видеть в просвете ацинусов этой железы. Таким образом, стало ясно, что АГ является промежуточным звеном между собственно синтезом секретируемого белка и выведением его из клетки. Также подробно процессы синтеза и выведения белков были изучены на других клетках (молочная железа, бокаловидные клетки кишечника, щитовидная железа и др.), и были исследованы морфологические особенности этого процесса. Синтезированный на рибосомах экспортируемый белок отделяется и накапливается внутри цистерн ЭР, по которым он транспортируется к зоне мембран АГ. Здесь от гладких участков ЭР отщепляются мелкие вакуоли, содержащие синтезированный белок, которые поступают в зону вакуолей в проксимальной части диктиосомы. В этом месте вакуоли могут сливаться друг с другом и с плоскими цис-цистернами диктиосомы. Таким образом происходит перенесение белкового продукта уже внутри полостей цистерн АГ.
По мере модификации белков в цистернах аппарата Гольджи, они с помощью мелких вакуолей переносятся от цистерн к цистерне в дистальную часть диктиосомы, пока не достигают трубчатой мембранной сети в транс-участке диктиосомы. В этом участке происходит отщепление мелких пузырьков, содержащих уже зрелый продукт. Цитоплазматическая поверхность таких пузырьков бывает сходна с поверхностью окаймленных пузырьков, которые наблюдаются при рецепторном пиноцитозе. Отделившиеся мелкие пузырьки сливаются друг с другом, образуя секреторные вакуоли. После этого секреторные вакуоли начинают двигаться к поверхности клетки, соприкасаются с плазматической мембраной, с которой сливаются их мембраны, и, таким образом, содержимое этих вакуолей оказывается за пределами клетки. Морфологически этот процесс экструзии (выбрасывания) напоминает пиноцитоз, только с обратной последовательностью стадий. Он носит название экзоцитоз.
Такое описание событий является только общей схемой участия аппарата Гольджи в секреторных процессах. дело усложняется тем, что одна и та же клетка может участвовать в синтезе многих выделяемых белков, может их друг от друга изолировать и направлять к клеточной поверхности или же в состав лизосом. В аппарате Гольджи происходит не просто “перекачка” продуктов из одной полости в другую, но и постепенно идет их “созревание”, модификация белков, которая заканчивается “сортировкой” продуктов, направляющихся или к лизосомам, или к плазматической мембране, или к секреторным вакуолям.
Модификация белков в аппарате Гольджи
В цис-зону аппарата Гольджи синтезированные в ЭР белки попадают после первичного гликозилирования и редукции там же нескольких сахаридных остатков. В конечном итоге все белки там имеют одинаковые олигосахаридные цепи, состоящие из двух молекул N-ацетилглюкозамина, шести молекул маннозы (рис. 182). В цис-цистернах начинается вторичная модификация олигосахаридных цепей и их сортировка на два класса. В результате олигосахариды на гидролитических ферментах, предназначенных для лизосом (богатые маннозой олгосахариды), фосфорилируются, а олигосахариды других белков, направляемых в секреторные гранулы, или к плазматической мембране, подвергаются сложным превращениям, теряя ряд сахаров и присоединяя галактозу, N-ацетилглюкозамин и сиаловые кислоты.
При этом возникает специальный комплекс олигосахаридов. Такие превращения олигосахаридов осуществляются с помощью ферментов - гликозилтрансфераз, входящих в состав мембран цистерн аппарата Гольджи. Так как каждая зона в диктиосомах имеет свой набор ферментов гликозилирования, то гликопротеиды как бы по эстафете переносятся из одного мембранного отсека (“этажа” в стопке цистерн диктиосомы) в другой и в каждом подвергаются специфическому воздействию ферментов. Так в цис-участке происходит фосфорилирование манноз в лизосомных ферментах и образуется особая маннозо-6-группировка, характерная для всех гидролитических ферментов, которые потом попадут в лизосомы.
В средней части диктиосом протекает вторичное гликозилирование секреторных белков: дополнительное удаление маннозы и присоединение N-ацетилглюкозамина. В транс-участке к олигосахаридной цепи присоединяются галактоза и сиаловые кислоты (рис. 183).
Эти данные были получены совершенно разными методами. С помощью дифференциального центрифугирования удалось получить раздельные более тяжелые (цис-) компоненты аппарата Гольджи и более легкие (транс-) и определить в них наличие гликозидаз и их продуктов. С другой стороны, используя моноклональные антитела к различным ферментам с помощью электронной микроскопии удалось их локализовать прямо на срезах клеток.
В ряде специализированных клеток в аппарате Гольджи происходит синтез собственно полисахаридов.
В аппарате Гольджи растительных клеток происходит синтез полисахаридов матрикса клеточной стенки (гемицеллюлозы, пектины). Кроме того, диктиосомы растительных клеток участвуют в синтезе и выделении слизей и муцинов, в состав которых входят также полисахариды. Синтез же основного каркасного полисахарида растительных клеточных стенок, целлюлозы, происходит как уже говорилось, на поверхности плазматической мембраны.
В аппарате Гольджи клеток животных происходит синтез длинных неразветвленных полисахаридных цепей глюкозаиногликанов. Один из них, гиалуроновая кислота, входящая в состав внеклеточного матрикса соединительной ткани, содержит несколько тысяч повторяющихся дисахаридных блоков. Многие глюкозаиногликаны ковалентно связаны с белками и образуют протеогликаны (мукопротеины). Такие полисахаридные цепи модифицируются в аппарате Гольджи и связываются с белками, которые в виде протеогликанов секретируются клетками. В аппарате Гольджи происходит также сульфатирование глюкозаиногликанов и некоторых белков.
Сортировка белков в аппарате Гольджи
Итак, через аппарат Гольджи проходит по крайней мере, три потока синтезированных клеткой нецитозольных белков: поток гидролитических ферментов в компартмент лизосом, поток выделяемых белков, которые накапливаются в секреторных вакуолях, и выделяются из клетки только по получении специальных сигналов, поток постоянно выделяемых секреторных белков. Следовательно, должен быть какой-то специальный механизм пространственного разделения этих разных белков и их путей следования.
В цис- и средних зонах диктиосом все эти белки идут вместе без разделения, они только раздельно модифицируются в зависимости от их олигосахаридных маркеров.
Собственно разделение белков, их сортировка, происходит в транс-участке аппарата Гольджи. Этот процесс не до конца расшифрован, но на примере сортировки лизосомных ферментов можно понять принцип отбора определенных белковых молекул (рис. 184).
Известно, что только белки-предшественники лизосомных гидролаз имеют специфическую олигосахаридную, а именно маннозную группу. В цис-цистернах эти группировки фосфорилируются и дальше вместе с другими белками переносятся от цистерны к цистерне, через среднюю зону в транс-участок. Мембраны транс-сети аппарата Гольджи содержат трансмембранный белок - рецептор (манноза-6-фосфатный рецептор или М-6-Ф-рецептор), который узнает фосфорилированные маннозные группировки олигосахаридной цепи лизосомных ферментов и связывается с ними. Это связывание происходит при нейтральных значениях рН внутри цистерн транс-сети. На мембранах эти М-6-Ф-рецепторные белки образуют кластеры, группы, которые концентрируются в зонах образования мелких пузырьков, покрытых клатрином. В транс-сети аппарата Гольджи происходит их отделение, отпочковывание и дальнейший перенос к эндосомам. Следовательно М-6-Ф-рецепторы, являясь трансмембранными белками, связываясь с лизосомными гидролазами, отделяют их, отсортировывают, от других белков (например, секреторных, нелизосомных) и концентрируют их в окаймленных пузырьках. Оторвавшись от транс-сети эти пузырьки быстро теряют шубу, сливаются с эндосомами, перенося свои лизосомные ферменты, связанные с мембранными рецепторами, в эту вакуоль. Как уже говорилось, внутри эндосом из-за активности протонного переносчика происходит закисление среды. Начиная с рН 6 лизосомные ферменты диссоциируют от М-6-Ф-рецепторов, активируются и начинают работать в полости эндолизосомы. Участки же мембран вместе с М-6-Ф-рецепторами возвращаются путем рециклизации мембранных пузырьков обратно в транс-сеть аппарата Гольджи.
Вероятнее всего, что та часть белков, которая накапливается в секреторных вакуолях и выводится из клетки после поступления сигнала (например нервного или гормонального) проходит такую же процедуру отбора, сортировки на рецепторах транс-цистерн аппарата Гольджи. Эти секреторные белки попадают сначала в мелкие вакуоли тоже одетые клатрином, которые затем сливаются друг с другом. В секреторных вакуолях часто происходит агрегация накопленных белков в виде плотных секреторных гранул. Это приводит к повышению концентрации белка в этих вакуолях примерно в 200 раз, по сравнению с его концентрацией в аппарате Гольджи. Затем эти белки по мере накопления в секреторных вакуолях выбрасываются из клетки путем экзоцитоза, поле получения клеткой соответствующего сигнала.
От аппарата Гольджи исходит и третий поток вакуолей, связанный с постоянной, конститутивной секрецией. Так фибробласты выделяют большое количество гликопротеидов и муцинов, входящих в основное вещество соединительной ткани. Многие клетки постоянно выделяют белки, способствующие связыванию их с субстратами, постоянно идет поток мембранных пузырьков к поверхности клетки, несущие элементы гликокаликса и мембранных гликопротеидов. Этот поток выделяемых клеткой компонентов не подлежит сортировке в рецепторной транс-системе аппарата Гольджи. Первичные вакуоли этого потока также отщепляются от мембран и относятся по своей структуре к окаймленным вакуолям, содержащим клатрин (рис. 185).
Заканчивая рассмотрение строения и работы такой сложной мембранной органеллы, как аппарат Гольджи, необходимо подчеркнуть, что несмотря на кажущуюся морфологическую однородность его компонентов, вакуоли и цистерны, на самом деле, это не просто скопище пузырьков, а стройная, динамичная сложно организованная, поляризованная система.
В АГ происходит не только транспорт везикул от ЕР к плазматической мембране. Существует ретроградный перенос везикул. Так от вторичных лизосом отщепляются вакуоли и возвращаются вместе с рецепторными белками в транс-АГ зону. Кроме того существует поток вакуолей от транс-зоны к цис-зоне АГ, а так же от цис-зоны к эндоплазматическому ретикулуму. В этих случаях вакуоли одеты белками COP I-комплекса. Считается, что таким путем возвращаются различные ферменты вторичного гликозилирования и рецепторные белки в составе мембран.
Эти особенности поведения транспортных везикул дали основу гипотезе о существовании двух типов транспорта компонентов АГ (рис. 186).
По одному из них, наиболее старому, в АГ существуют стабильные мембранные компоненты, к которым от ЭР эстафетно переносятся вещества с помощью транспортных вакуолей. По альтернативной модели АГ является динамическим производным ЭР: отщепившиеся от ЭР мембранные вакуоли сливаются друг с другом в новую цис-цистерну, которая затем продвигается через всю зону АГ и в конце распадается на транспортные везикулы. По этой модели ретроградные COP I везикулы возвращают постоянные белки АГ в более молодые цистерны. Таким образом предполагается, что переходная зона ЭР представляет собой “родильный дом” для аппарата Гольджи.
studfiles.net
Комплекс (аппарат) Гольджи, строение и функции
Аппарат Гольджи – одномембранная, микроскопическая органелла эукариотической клетки, которая предназначена для завершения процессов синтеза клетки и обеспечивает вывод образовавшихся веществ.
Исследование структурных компонентов комплекса Гольжи началось еще в 1898 итальянским ученым-гистологом Камилло Гольджи, в честь него органелла и была названа. Изучение органоида проходило впервые в составе нервной клетки.
 Внешний вид комплекса Гольджи
Внешний вид комплекса ГольджиСтроение комплекса Гольджи
В пластинчатом комплексе (аппарат Гольджи) имеется три части:
- Цис-цистерна — находится вблизи ядра, постоянно взаимодействует с гранулярной эндоплазматической сетью;
- медиал-цистерна или промежуточная часть;
- транс-цистерна — отдаленная от ядра, дает трубчатые разветвления, формируя транс-сеть Гольджи.
Пластинчатый комплекс в клетках разной природы и даже на различных этапах дифференцировки одной клетки, иногда имеет отличительные черты в строении.
 Строение аппарата Гольджи
Строение аппарата ГольджиХарактерные признаки аппарата Гольджи
Имеет вид стопки, которая состоит от трех до восьми цистерн, толщиной около 25 нм, они уплощены в центральной части и расширяются в направлении к периферии, напоминают стопку перевернутых тарелок. Поверхности цистерн примыкают друг к другу очень плотно. От периферической части отпочковываются небольшие мембранные пузырьки.
Клетки человека имеют одну, реже пару стопок, а клетки растений могут содержать несколько таких образований. Совокупность цистерн (одна стопка) совместно с окружающими ее пузырьками называется диктиосомой. Несколько диктиосом могут связываться между собой, формируя сеть.
Полярность – наличие цис-стороны, направленной к ЭПС и ядру, где происходит слияние везикул, и транс-стороны, устремленной к клеточной оболочке (это особенность хорошо прослеживается в клетках секретирующих органов).
Асимметричность – сторона расположенная ближе к ядру клетки (проксимальный полюс) вмещает «незрелые» белки, к ней постоянно присоединяются везикулы, отсоединившиеся от ЭПС, транс-сторона (дистальный, зрелый полюс) содержит уже модифицированные белки.
При разрушении чужеродными агентами пластинчатого комплекса, происходит разделение аппарата Гольджи на отдельные части, но его основные функции при этом сохраняются. После возобновления системы микротрубочек, которые были хаотично разбросаны в цитоплазме, части аппарата собираются, и снова превращаются в нормально функционирующий пластинчатый комплекс. Физиологическое разделение происходит и в обычных условиях жизнедеятельности клеток, во время непрямого деления.
ЭПС и комплекс Гольджи
ЭПС – это часть комплекса Гольджи?
Однозначно нет. Эндоплазматическая сеть – это самостоятельная мембранная органелла, которая построена из системы замкнутых канальцев, цистерн, сформированных непрерывной мембраной. Основная функция – синтез белков, с помощью рибосом, размещенных на поверхности гранулярной ЭПС.
Существует ряд сходных признаков между ЭПС и аппаратом Гольджи:
- Это внутриклеточные образования, отграниченные от цитоплазмы мембраной;
- отделяют мембранные пузырьки, которые наполнены органическими продуктами синтеза;
- вместе формируют единую синтезирующую систему;
- в секретирующих клетках имеют наибольшие размеры и высокий уровень развития.
Чем образованы стенки эндоплазматической сети и комплекса Гольджи?
Стенки ЭПС и аппарата Гольджи представлены в виде однослойной мембраны. Эти органеллы вместе с лизосомами, пероксисомами и митохондриями объединены в группу мембранных органоидов.
Что происходит в комплексе Гольджи с гормонами и ферментами?
За синтез гормонов отвечает эндоплазматическая сеть, на поверхности ее мембраны идет производство гормональных веществ. В комплекс Гольджи поступают синтезированные гормоны, здесь они накапливаются, затем идет переработка и выведение их наружу. Поэтому в клетках эндокринных органов встречаются комплексы больших размеров (до 10 мкм).
Функции комплекса Гольджи
Протеолиз белковых веществ, что приводит к активации белков, так проинсулин переходит в инсулин.
Обеспечивает транспорт из клетки продуктов синтеза ЭПС.
Самой важной функцией комплекса Гольджи считают выведение из клетки продуктов синтеза, поэтому его еще называют транспортным аппаратом клетки.
Синтез полисахаридов, таких как пектин, гемицеллюлоза, которые входят в состав мембран растительных клеток, образование гликозаминогликанов, одного из составляющих межклеточной жидкости.
В цистернах пластинчатого комплекса идет созревание белковых веществ, необходимых для секреции, трансмембранных протеинов клеточной мембраны, ферментов лизосом и др. В процессе созревания белки постепенно перемещаются по отделам органоида, в которых завершается их формирование и происходит гликозилирование и фосфорилирование.
Формирование липоптротеидных веществ. Синтез и накопление слизистых веществ (муцина). Образование гликолипидов, которые входят в состав мембранного гликокаликса.
Передает белки в трех направлениях: к лизосомам (перенос контролируется ферментом – маннозой- 6-фосфат), к мембранам или внутриклеточной среде, и к межклеточному пространству.
Вместе с зернистой ЭПС образует лизосомы, путем слияния отпочковавшихся везикул с автолитическими ферментами.
Экзоцитозный перенос – везикула, подойдя к мембране, встраивается в нее и оставляет свое содержимое с наружной стороны клетки.
Сводная таблица функций комплекса Гольджи
| Цис-цистерна | Захват синтезированных ЭПС белков, мембранных липидов |
| Срединные цистерны | Посттрансляционные модификации связанные с переносом ацетилглюкозамина. |
| Транс-цистерна | Завершается гликозилирование, присоединение галактозы и сиаловой кислоты, идет сортировка веществ для дальнейшего транспорта из клетки. |
| Пузырьки | Отвечают за перенос липидов, белков в аппарат Гольджи и между цистернами, а также за выведение продуктов синтеза. |
animals-world.ru
Эндоплазматическая сеть, аппарат Гольджи, лизосомы и вакуоли образуют единую вакуолярную сеть клетки, отдельные элементы которой могут переходить друг в друга
План урока №6
ТЕМА: «Органоиды клетки».
ЦЕЛИ:
1. Образовательная:
- дать представление о строении и роли органоидов в жизни клетки.
2. Развивающая:
- развить логическое мышление, умения устанавливать причинно-следственные связи
3.Воспитательная:
- формировать естественное отношение к учебному процессу
Ход урока:
I Организационный момент
1. Ознакомление студентов с темой и целью урока.
2. Перед студентами ставится ряд заданий, которые необходимо выполнить в процессе урока:
- иметь представление об органоидах цитоплазмы, знать их строение и функции
- уметь связать функции органоидов клетки с физиологическими, процессами протекающими в ней
II Основная часть
1. Проверка домашнего задания
- что такое клетка, назвать главные части клетки
- клеточная оболочка и ее основные функции
- фагоцитоз, пиноцитоз
- цитоплазма и ее функции
- строение и роль ядра
- назовите главные особенности строения растительной клетки
2. Объяснение нового материала
а) Эндоплазматическая сеть, ее функции в клетке
б) Рибосомы и их роль
в) Митохондрии – строение и функции
г) Строение и функции клеточного центра
д) Функции комплекса Гольджи
е) Роль лизосом в клеточном пищеварении
3. Закрепление нового материала
- в чем различие между гладкими и шероховатыми мембранами ЭПС, назовите функции ЭПС
- из чего состоят и где расположены рибосомы
- почему митохондрии называют энергетическими станциями клеток
- что представляет собой клеточный центр и органоиды движения
- значение аппарата Гольджи в жизни клетки
III Подведение итогов урока
IV Домашнее задание
1. Записи в тетрадях.
2. Учебник Ю.И.Полянского «Общая биология» (стр. 132-139)
3. Учебник В.Б.Захаров, С.Т.Мамонтов «Биология» (стр.147-154)
4. Подготовка докладов и сообщений: «Открытие вирусов в организме»
«Структура вирусов, генетический аппарат вирусов, оболочка вирусов, жизненный цикл вируса».
5. Подготовка к тестированию по теме: «Строение клетки»
Занятие №6
Тема: «Органоиды клетки».
1. Одномембранные органоиды.
2. Двумембранные органоиды.
3. Немебранные органоиды.
Различают:
1.Мембранные органоиды — имеющие мембранное строение, причем они могут быть
а)одномембранными: эндоплазматическая сеть, аппарат Гольджи, лизосомы, вакуоли
б) двумембранными: митохондрии, пластиды, ядро
2.Не имеющие мембранного строения—хромосомы, рибосомы, клеточный центр, центриоли, реснички и жгутики, микротрубочки
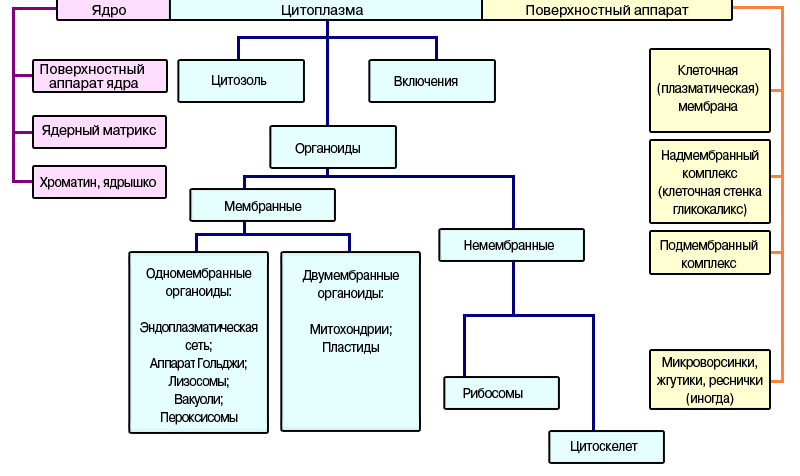
Одномембранные:
Эндоплазматическая сеть (ЭПС) — одномембранный органоид, представляет собой ажурную конструкцию из соединенных полостей, канальцев и трубочек,пронизывающих цитоплазму клетки. Она образована мембраной, сходной по строению с ПМ. Трубочки и полости ЭПС могут занимать до 50% объема клетки и нигде не обрываются и не открываются в цитоплазму. Мембраны с одной стороны связаны с цитоплазматической мембраной, с другой — с наружной ядерной мембраной.
Различают два вида мембран ЭПС:
1)Шероховатую– содержит на своей поверхности рибосомы. Именно здесь синтезируется большинство белков. Шероховатая сеть лучше развита в тех клетках, которые синтезируют белки для нужд всего организма (например, белковые гормоны)
2) Гладкую - мембраны которой рибосом не несут. На поверхности гладкой ЭПС идет синтез углеводов и липидов. Развита в тех клетках, которые синтезируют, к примеру, сахара и липиды. В гладкой ЭПС, кроме того, накапливаются ионы кальция — важные регуляторы всех функций клеток и целого организма.
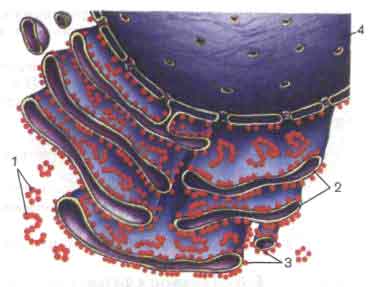
1 - свободные рибосомы; 2 - полости;3 - рибосомы, прикрепленные к мембранам;4 - ядерная оболочка.
Функции:
1) транспорт веществ из одной части клетки в другую,
2) разделение цитоплазмы клетки на компраменты («отсеки»),
3) синтез углеводов и липидов (гладкая ЭПС),
4) синтез белка (шероховатая ЭПС),
5) место образования аппарата Гольджи.
Аппарат Гольджи, или комплекс Гольджи, — одномембранный органоид. Описан в 1889 году. Аппарат Гольджи обычно расположен около клеточного ядра (в животных клетках часто вблизи клеточного центра). При специальной окраске различим в оптическом микроскопе (имеет вид сетчатой структуры). Состоит из:
-уплощенных мешочков «цистерн» с расширенными краями – имеют вид дискообразных полостей, расположенных часто группами по 13–15 (диктиосомы). Каждая стопка обычно состоит из 4-х–6-ти «цистерн», является структурно-функциональной единицей аппарата Гольджи и называется диктиосомой. Число диктиосом в клетке колеблется от одной до нескольких сотен;
- крупных вакуолей (пузырьки Гольджи) – образуются в результате расширения цистерн; мелких вакуолей – отшнуровываются от краев цистерн. Их число доходит до нескольких тысяч.
Структура и функция комплекса Гольджи 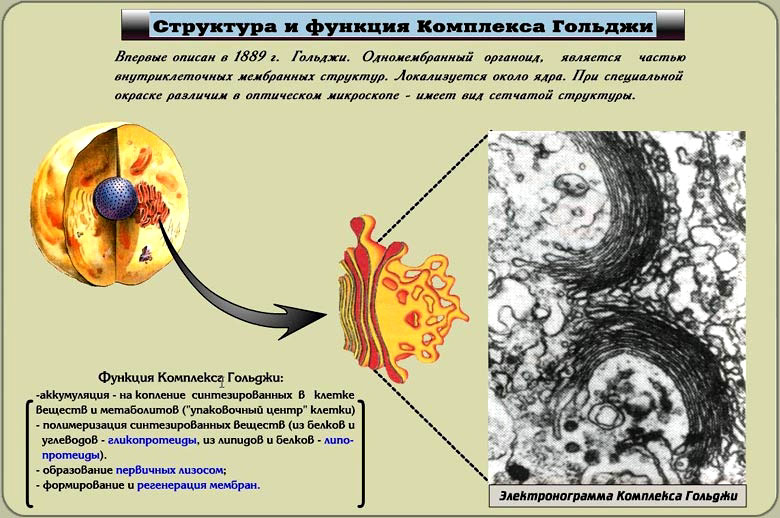
Функции:
1) накопление белков, липидов, углеводов,
2) модификация поступивших органических веществ,
3) «упаковка» в мембранные пузырьки белков, липидов, углеводов,
4) секреция белков, липидов, углеводов,
5) синтез углеводов и липидов,
6) место образования лизосом.
Секреторная функция является важнейшей, поэтому аппарат Гольджи хорошо развит в секреторных клетках.
Лизосомы — от “лизио” – растворяю и “сома” – тело - одномембранные органоиды. Представляют собой мелкие пузырьки (диаметр от 0,2 до 0,8 мкм), Характерны для клеток животных, грибов, в растениях не выявлены. Различают 4 вида лизосом:
- первичная лизосома – содержит неактивные ферменты, синтезированные рибосомами, накопленными в ЭПС и поступившими в комплекс Гольджи, который упаковывает их в мембранный пузырек.
- вторичная лизосома – гетерофагосома или пищеварительная вакуоль, возникает как результат соединения первичной лизосомы с поглощенным клеткой (путем фаго, и пиноцитоза) чужеродным материалом или собственными компонентами клетки, предназначенными для расщепления. Поглощенный материал постепенно переваривается под действием гидролаз поступивших в фагосому, переваренные вещества проходят через мембрану фагосомы и включаются в состав клетки.
- остаточные тельца – содержат непереваренные вторичными лизосомами питательные вещества. У простейших остаточные тельца выделяются во внешнюю среду. В других случаях они могут длительное время сохраняться в клетке и вызывать различные патологические процессы (у человека известно около 12 врожденных заболеваний, при которых отмечается дисфункция лизосом).
- цитолизосома – образуется при соединении первичной лизосомы с компонентами самой клетки (например, митохондрий или участков ЭПС). Они образуются в ходе различных физиологических (регенерация) и патологических процессов.
Свойства лизосом:
- образуются в комплексе Гольджи.
- содержатся в клетке от 10 до 100 и более.
- содержат около 60 гидролаз (класс ферментов, катализирующих реакции расщепления различных веществ: белков, жиров, углеводов, при участии молекул воды).
Структура и функции лизосом.
мембрана
ферменты
Функции:
1) внутриклеточное переваривание органических веществ,
2) уничтожение ненужных клеточных и неклеточных структур,
3) участие в процессах реорганизации клеток.
Вакуоли — одномембранные органоиды, представляют собой «емкости», заполненные водными растворами органических и неорганических веществ. В образовании вакуолей принимают участие ЭПС и аппарат Гольджи. Молодые растительные клетки содержат много мелких вакуолей, которые затем по мере роста и дифференцировки клетки сливаются друг с другом и образуют одну большую центральную вакуоль. Центральная вакуоль может занимать до 95% объема зрелой клетки, ядро и органоиды оттесняются при этом к клеточной оболочке. Мембрана, ограничивающая растительную вакуоль, называется тонопластом. Жидкость, заполняющая растительную вакуоль, называется клеточным соком. В состав клеточного сока входят водорастворимые органические и неорганические соли, моносахариды, дисахариды, аминокислоты, конечные или токсические продукты обмена веществ (гликозиды, алкалоиды), некоторые пигменты (антоцианы).
В животных клетках имеются мелкие пищеварительные и автофагические вакуоли, относящиеся к группе вторичных лизосом и содержащие гидролитические ферменты. У одноклеточных животных есть еще сократительные вакуоли, выполняющие функцию осморегуляции и выделения.
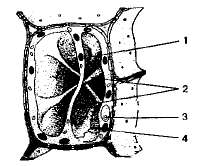
1 — вакуоль; 2 — цитопяаз-матические тяжи; 3 — ядро; 4 — хлоропласты.
Функции:
1) накопление и хранение воды,
2) регуляция водно-солевого обмена,
3) поддержание тургорного давления,
4) накопление водорастворимых метаболитов, запасных питательных веществ,
5) окрашивание цветов и плодов и привлечение тем самым опылителей и распространителей семян
Эндоплазматическая сеть, аппарат Гольджи, лизосомы и вакуоли образуют единую вакуолярную сеть клетки, отдельные элементы которой могут переходить друг в друга.
Двумембранные:
Митохондрии«митос»- нить, «хондрион» -зерно–энергетические органоиды их еще называют «силовыми станциями» клеток. Форма митохондрий различна, они м.б. овальными, палочковидными, нитевидными. Число митохондрий зависит от функциональной активности клетки и может достигать десятка тысяч в летательных мышцах насекомых.Оболочка митохондрии состоит из двух мембран – наружной – гладкой (не образующую никаких складок) и внутренней- образует многочисленные складки – кристы (лат. «криста» - гребень, вырост). Внутри митохондрий находятся РНК, ДНК и рибосомы отличающиеся от цитоплазматических. Между кристами находится вязкая белоксодержащая масса - матрикс,в нем содержатся различные ферменты. С помощью этих ферментов происходит преобразование энергии питательных веществ в энергию АТФ - необходимую для жизнедеятельности клетки и организма в целом.
Согласно одной из гипотез (теория симбиогенеза) митохондрии произошли от древних свободноживущих аэробных прокариотических организмов, которые, случайно проникнув в клетку-хозяина, затем образовали с ней взаимовыгодный симбиотический комплекс. В пользу этой гипотезы свидетельствуют следующие данные. Во-первых, митохондриальная ДНК имеет такие же особенности строения как и ДНК современных бактерий (замкнута в кольцо, не связана с белками). Во-вторых, митохондриальные рибосомы и рибосомы бактерий относятся к одному типу — 70S-типу. В-третьих, механизм деления митохондрий сходен с таковым бактерий. В-четвертых, синтез митохондриальных и бактериальных белков подавляется одинаковыми антибиотиками.
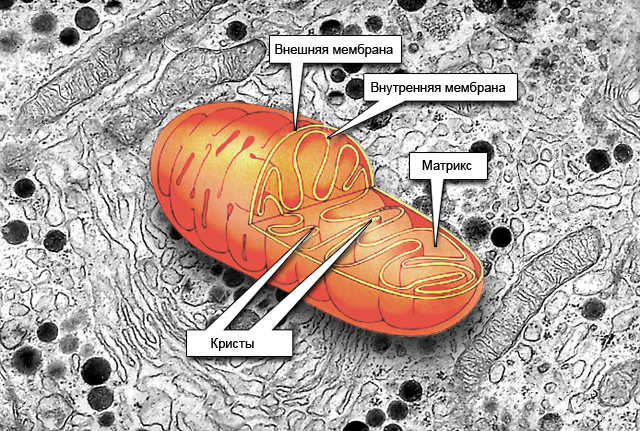
Основная функция–1) синтезАТФ;
2)кислородное расщепление органических веществ.
Пластиды
Пластиды характерны только для растительных клеток. Различают три основных типа пластид: лейкопласты — бесцветные пластиды в клетках неокрашенных частей растений, хромопласты — окрашенные пластиды обычно желтого, красного и оранжевого цветов, хлоропласты — зеленые пластиды. Строение пластид: 1 — наружная мембрана; 2 — внутренняя мембрана; 3 — строма; 4 — тилакоид; 5 — грана.
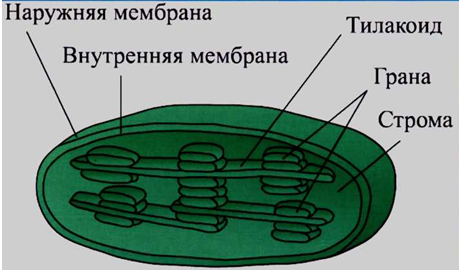
Хлоропласты. В клетках высших растений хлоропласты имеют форму двояковыпуклой линзы. Длина хлоропластов колеблется в пределах от 5 до 10 мкм, диаметр — от 2 до 4 мкм. Хлоропласты ограничены двумя мембранами. Наружная мембрана (1) гладкая, внутренняя (2) имеет сложную складчатую структуру. Наименьшая складка называется тилакоидом (4). Группа тилакоидов, уложенных наподобие стопки монет, называется граной (5). В хлоропласте содержится в среднем 40–60 гран, расположенных в шахматном порядке. Граны связываются друг с другом уплощенными каналами — ламеллами (6). В мембраны тилакоидов встроены фотосинтетические пигменты и ферменты, обеспечивающие синтез АТФ. Главным фотосинтетическим пигментом является хлорофилл, который и обусловливает зеленый цвет хлоропластов.
Внутреннее пространство хлоропластов заполнено стромой (3). В строме имеются кольцевая «голая» ДНК, рибосомы 70S-типа, ферменты цикла Кальвина, зерна крахмала (7). Внутри каждого тилакоида находится протонный резервуар, происходит накопление Н+. Хлоропласты, также как митохондрии, способны к автономному размножению путем деления надвое. Они содержатся в клетках зеленых частей высших растений, особенно много хлоропластов в листьях и зеленых плодах. Хлоропласты низших растений называют хроматофорами.
Функция хлоропластов: фотосинтез. Полагают, что хлоропласты произошли от древних эндосимбиотических цианобактерий (теория симбиогенеза). Основанием для такого предположения является сходство хлоропластов и современных бактерий по ряду признаков (кольцевая, «голая» ДНК, рибосомы 70S-типа, способ размножения).
Лейкопласты. Форма варьирует (шаровидные, округлые, чашевидные и др.). Лейкопласты ограничены двумя мембранами. Наружная мембрана гладкая, внутренняя образует малочисленные тилакоиды. В строме имеются кольцевая «голая» ДНК, рибосомы 70S-типа, ферменты синтеза и гидролиза запасных питательных веществ. Пигменты отсутствуют. Особенно много лейкопластов имеют клетки подземных органов растения (корни, клубни, корневища и др.).
Функция лейкопластов: синтез, накопление и хранение запасных питательных веществ. Амилопласты — лейкопласты, которые синтезируют и накапливают крахмал, элайопласты — масла, протеинопласты — белки. В одном и том же лейкопласте могут накапливаться разные вещества.
Хромопласты.Ограничены двумя мембранами. Наружная мембрана гладкая, внутренняя или также гладкая, или образует единичные тилакоиды. В строме имеются кольцевая ДНК и пигменты — каротиноиды, придающие хромопластам желтую, красную или оранжевую окраску. Форма накопления пигментов различная: в виде кристаллов, растворены в липидных каплях (8) и др. Содержатся в клетках зрелых плодов, лепестков, осенних листьев, редко — корнеплодов. Хромопласты считаются конечной стадией развития пластид.
Функция хромопластов: окрашивание цветов и плодов и тем самым привлечение опылителей и распространителей семян.
Все виды пластид могут образовываться из пропластид. Пропластиды — мелкие органоиды, содержащиеся в меристематических тканях. Поскольку пластиды имеют общее происхождение, между ними возможны взаимопревращения. Лейкопласты могут превращаться в хлоропласты (позеленение клубней картофеля на свету), хлоропласты — в хромопласты (пожелтение листьев и покраснение плодов). Превращение хромопластов в лейкопласты или хлоропласты считается невозможным.
Немембранные:
Рибосомы. Это мелкие органеллы (диаметром порядка 20 нм), встречающиеся в клетках всех организмов. Рибосомы располагаются либо на мембранах шероховатой эндоплазматической сети, либо свободно лежат в цитоплазме. И в тех и других синтезируются белки. В одной клетке содержится много тысяч рибосом.
Строение: Каждая рибосома состоит из двух неодинаковых по размерам частиц, малой и большой. Образуется в ядрышке из ДНК. В состав рибосом входят белки и рибосомальные РНК (р- РНК). Молекулы рРНК составляют 90% массы рибосомы и образуют ее структурный каркас. Большинство белков специфически связано с определенными участками рРНК. Некоторые белки входят в состав рибосом только во время биосинтеза белка.
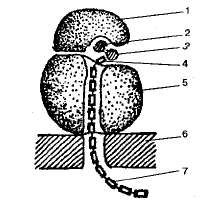 Схема строения рибосомы:
Схема строения рибосомы:
1 — малая субъединица; 2 — иРНК; 3 — тРНК; 4 — аминокислота; 5 — большая субьединица; б — мембрана эндоплазматической сети; 7 — синтезируемая полипептид-ная цепь.
Функции: обеспечивает синтез белка (сборку белковой молекулы из аминокислот).
Синтез белка - сложный процесс, который осуществляется не одной рибосомой, а целой группой, включающей до нескольких десятков объединенных рибосом. Такую группу рибосом называют полисомой.
Синтезированные белки сначала накапливаются в каналах и полостях эндоплазматической сети, а затем транспортируются к органоидам и участкам клетки, где они потребляются.
studopedya.ru
Гольджи аппарат: введение
Гольджи аппарат: введение
Аппарат Гольджи является компонентом всех эукариотических клеток (практически единственное исключение - эритроциты млекопитающих). Он представляет собой важнейшую мембранную органеллу, управляющую процессами внутриклеточного транспорта .
Аппарат Гольджи состоит из окруженных мембранами полостей, уложенных в стопку, в которых происходит сортировка и упаковка поступивших макромолекул ( рис. 26 ). Вещества, необходимые клетке для внутреннего потребления, накапливаются в мелких мембранных пузырьках (50 нм), отпочковывающихся от полостей аппарата Гольджи, и разносятся по цитоплазме. Вещества, произведенные клеткой "на экспорт", получают в аппарате Гольджи "удостоверение" (метку) в виде присоединенных к ним полисахаридов и в таком виде выводятся из клетки. Наиболее крупные (до 10 мкм) аппараты Гольджи обнаружены в клетках желез внутренней секреции. В них производятся гормоны, которые попадают в кровяное русло и разносятся ко всем клеткам тела. "Удостоверение", полученное в аппарате Гольджи, позволяет гормону дойти до адресата, а не быть уничтоженным по дороге за "контрабанду". Одна из функций аппарата Гольджи - участие в образовании лизосом . Лизосомы участвуют в расщеплении "старых" частей клетки, целых клеток и отдельных органов.
Основными функциями аппарата Гольджи являются модификация, накопление, сортировка и направление различных веществ в соответствующие внутриклеточные компартменты, а также за пределы клетки. Он состоит из набора окруженных мембраной уплощенных цистерн , напоминающих стопку тарелок. Каждая стопка Гольджи (у растений называемая диктиосомой ) обычно содержит от четырех до шести цистерн, имеющих, как правило, диаметр около 1мкм ( рис. 8-36 ). Число стопок Гольджи в клетке в значительной степени зависит от ее типа: некоторые клетки содержат одну большую стопку, тогда как в других имеются сотни очень маленьких стопок.
Со стопками Гольджи всегда ассоциирована масса мелких (диаметром приблизительно 60 нм) ограниченных мембраной пузырьков. Полагают, что эти пузырьки ( пузырьки Гольджи ) переносят белки и липиды в аппарат Гольджи, транспортируют их из него и между остальными цистернами. Многие пузырьки являются окаймленными и покрыты клатрином или другим специфическим белком. Часто можно видеть, как такие окаймленные пузырьки отшнуровываются от цистерн Гольджи.
Аппарат Гольджи имеет две разные стороны: формирующуюся, или цис-сторону и зрелую, или транс-сторону . Цис-сторона тесно связана с переходными элементами ЭР; транс-сторона расширяется, образуя трубчатый ретикулум, называемый транс-сетью Гольджи . Белки и липиды в составе небольших пузырьков попадают в стопку Гольджи с цис-стороны, а покидают ее, направляясь в различные компартменты, вместе с пузырьками, образующимися на транс-стороне. Переходя из одной стопки Гольджи в другую, эти молекулы претерпевают последовательные серии модификаций.
Несмотря на то, что в строении и механизмах функционирования аппарата Гольджи остается еще много неясного, очевидно, что он выполняет чрезвычайно многообразные и важные для клетки функции. Наиболее изученные из них состоят в транспорте и химической модификации секретируемых продуктов.
Хорошо развитый аппарат Гольджи присутствует не только в секреторных, но и практически во всех клетках эукариотических организмов. Это не удивительно, т.к. любая клетка должна постоянно регенерировать и обновлять свою мембрану, поддерживать специфичность своих органелл, образовывать лизосомы и т.п.
В последнее время интерес исследователей к аппарату Гольджи резко возрос, и в результате быстро накапливаются все новые и новые данные об этой интереснейшей органелле и ее участии в процессах внутриклеточного мембранного транспорта.Кому принадлежит честь открытия аппарата Гольджи, до сих пор неизвестно. Впервые похожую структуру описал La Valette St. George в 1865-1867 годах. Впоследствии многие ученые отмечали сходство этой структуры с органеллой, описанной Камилло Гольджи в 1898 году. Однако имеются сомнения в том, что La Valette St. George действительно ясно видел ту структуру, которую описывал. Независимо от Гольджи " диктиосому " (как стали называть новую органеллу) наблюдали Platner (1889), Murray (1898) и, вероятно, немало других исследователей, принимая во внимание разнообразие изучавшихся тогда тканей и клеток, а также многочисленность применявшихся способов фиксации и окраски препаратов. Тем не менее именно Гольджи первым создал метод, который позволил безошибочно выделить новую структуру на фоне других клеточных органелл и описать некоторые детали ее строения и местонахождения в клетке. Метод Гольджи состоял в фиксации ткани раствором бихромата натрия с последующей окраской азотнокислым серебром. С помощью этого метода Гольджи обнаружил в некоторых нервных клетках неизвестную ему ранее структуру, которую он назвал "внутренней сетью". Впоследствии, с усовершенствованием методов, было доказано существование этой органеллы в клетках других тканей.
Ссылки:
medbiol.ru
определение, строение, функции и история открытия
Что такое комплекс Гольджи?
Аппарат Гольджи, также называемый комплексом Гольджи — органелла встречающая, как в клетках растений, так и животных, и обычно состоит из совокупности чашеобразных отделов с мембраной, называемых цистернами, которые выглядят как стопка сдутых воздушных шаров.
Однако у некоторых одноклеточных жгутиковых имеется 60 цистерн, формирующих аппарат Гольджи. Точно так же количество стопок комплекса Гольджи в клетке изменяется в зависимости от ее функций. Клетки животных, как правило, содержат от 10 до 20 стопок на одну клетку, объединенных в один комплекс трубчатыми соединениями между цистернами. Аппарат Гольджи обычно расположен близко к ядру клетки.
История открытия
Из-за относительно больших размеров комплекс Гольджи был одной из первых наблюдаемых органелл в клетках. В 1897 году итальянский врач по имени Камилло Гольджи, изучающий нервную систему, использовал новую технологию окрашивания, которую сам же разработал (и которая актуальна в наши дни). Благодаря новому методу, ученый смог разглядеть клеточную структуру и назвал ее внутренним ретикулярным аппаратом.
Вскоре после того, как он публично объявил о своем открытии в 1898 году, структура была названа в его честь, становясь универсально известной как аппарат Гольджи. Тем не менее, многие ученые того времени не верили, что Гольджи наблюдал настоящую органеллу клетки, и списывали открытие ученного на визуальное искажение, вызванное окрашиванием. Изобретение электронного микроскопа в двадцатом веке окончательно подтвердило, что аппарат Гольджи является клеточной органеллой.
Строение
У большинства эукариот аппарат Гольджи формируется из стопок мешочков, состоящих из двух основных отделов: цис-отдела и транс-отдела. Цис-отдел представляет собой комплекс сплюснутых мембранных дисков, известных как цистерны, происходящие из везикулярных кластеров, которые устремляются из эндоплазматического ретикулума.
Клетки млекопитающих обычно содержат от 40 до 100 стопок. Как правило, от в каждая стопка включает от 4 до 8 цистерн. Однако у некоторых протистов наблюдается около 60 цистерн. Этот набор цистерн разбит на цис, медиальные и транс-отделы. Транс-отдел представляет собой конечную цистернальную структуру, из которой белки упаковываются в везикулы, предназначенные для лизосом, секреторных везикул или клеточной поверхности.
Функции
Аппарат Гольджи часто считается отделом распределения и доставки химических веществ клетки. Он модифицирует белки и липиды (жиры), которые продуцируются в эндоплазматическом ретикулуме, и готовит их для экспорта за пределы клетки или для транспортировки в другие места внутри клетке. Белки и липиды, построенные в гладком и шероховатом эндоплазматическом ретикулуме, укладываются в крошечные пузырьковые везикулы, которые движутся через цитоплазму, пока не достигнут комплекса Гольджи.
Везикулы сливаются с мембранами Гольджи и высвобождают, содержащиеся внутри молекулы в органеллу. Оказавшись внутри, соединения дополнительно обрабатываются с помощью аппарата Гольджи, а затем направляются в везикуле к месту назначения внутри или вне клетки. Экспортируемые продукты представляют собой секреции белков или гликопротеинов, которые являются частью функции клетки в организме. Другие вещества возвращаются в эндоплазматический ретикулум или могут созревать, чтобы впоследствии стать лизосомами.
Модификации молекул, которые осуществляются в комплексе Гольджи, происходят упорядоченным образом. Каждая цистерна имеет два основных отдела: цис-отдел — это конец органеллы, где вещества поступают из эндоплазматического ретикулума для обработки, а также транс-отдел, где они выходят в форме меньших отдельных везикул. Следовательно, цис-отдел расположен вблизи эндоплазматического ретикулума, откуда поступает большая часть веществ, а транс-отдел расположен вблизи плазматической мембраны клетки, куда отправляются многие из веществ, модифицирующиеся в аппарате Гольджи.
Химический состав каждого отдела, а также ферменты, содержащиеся в люменах (внутренних открытых пространствах цистерн) между отделами, являются отличительными. Белки, углеводы, фосфолипиды и другие молекулы, образующиеся в эндоплазматическом ретикулуме, переносятся на аппарат Гольджи, чтобы подвергнутся биохимическому модифицированию при переходе от цис к транс-отделам комплекса. Ферменты, присутствующие в люмене Гольджи, модифицируют углеводную часть гликопротеинов путем добавления или вычитания отдельных мономеров сахара. Кроме того, аппарат Гольджи сам по себе производит самые разнообразные макромолекулы, включая полисахариды.
Комплекс Гольджи в растительных клетках продуцирует пектины и другие полисахариды, необходимые для структуры растений и обмена веществ. Продукты, экспортируемые аппаратом Гольджи через транс-отдел, в конечном итоге сливаются с плазматической мембраной клетки. Среди наиболее важных функций комплекса — сортировка большого количества макромолекул, продуцируемых клеткой, и их транспортировка в необходимые пункты назначения. Специализированные молекулярные идентификационные метки или метки, такие как фосфатные группы, добавляются ферментами Гольджи, чтобы помочь в этом процессе сортировки.
← Подписывайтесь на наши аккаунты в соц.сетях, чтобы не пропустить самую интересную информацию!natworld.info

|
|
..:::Счетчики:::.. |
|
|
|
|
|
|
|
|